- POLYMORPHISME BIOCHIMIQUE DES ÊTRES VIVANTS
- POLYMORPHISME BIOCHIMIQUE DES ÊTRES VIVANTSLes populations naturelles d’organismes, animaux ou végétaux, présentent une certaine hétérogénéité qui correspond soit à des formes saisonnières, soit à une variabilité géographique, soit à une variabilité génétique qui peut être continue (exprimée généralement sous la forme d’une courbe en cloche et représentant la variation appelée quantitative en génétique des populations) ou discontinue. C’est à cette variation discontinue intraspécifique que l’on a donné le nom de polymorphisme, défini avec précision par E. B. Ford (1940) comme «la présence simultanée et dans le même biotope de deux ou plusieurs formes discontinues, ou morphes ou phases, appartenant à une même espèce et existant en proportions telles que la plus rare d’entre elles ne peut être maintenue au sein de l’espèce par la pression de mutation récurrente». L’origine de ces morphes est à rechercher dans des phénomènes mutationnels qui auraient pu se produire à un quelconque niveau du génome (ségrégations d’allèles à un locus génétique, ségrégation de fragments chromosomiques ou de chromosomes entiers). Les morphes ne sont donc que des phénotypes représentant l’action d’un ou de plusieurs gènes, ou encore d’un ensemble considérable de gènes. Ainsi, des différences génétiques inscrites dans le génotype de chaque individu produisent-elles des différences phénotypiques au niveau de la population. Ces différences individuelles se traduisent par la présence de variantes, décelables par diverses techniques, allant de la simple inspection visuelle à l’analyse des chromosomes (caryotype), à des tests physiologiques ou biochimiques ou encore à des tests génétiques plus ou moins compliqués. Certaines des différences qui font partie du polymorphisme sont donc parfaitement aisées à mettre à évidence, alors que d’autres sont entièrement cachées par des mécanismes physiologiques et génétiques, et nécessitent des méthodes particulières pour pouvoir être analysées.Le rôle du polymorphisme dans les populations naturelles d’organismes apparaît immense, puisque c’est la matière première indispensable aux mécanismes de sélection naturelle. À long terme, cette variabilité permet de concevoir comment des effets sélectifs agissant sur des populations géographiquement isolées les unes des autres peuvent aboutir à la différenciation des espèces, ou spéciation [cf. SPÉCIATION]. Si, dans le strict contexte écologique du lieu et du moment où elle se produit, une mutation augmente la valeur sélective de l’individu qui la recèle, elle peut se répandre dans la population et augmenter de fréquence. En jargon de génétique évolutive, on dit que cette mutation est favorable pour l’organisme ou qu’elle est favorisée par la sélection naturelle. Dans un tel cas, il y aura deux formes génétiques dans la population, formes dont la fréquence va dépendre des conditions écologiques, car, si le mutant est favorable à un temps donné t , il n’est pas certain qu’il le soit encore à un temps t + 1. Si la mutation est délétère au moment de son apparition dans le génome et diminue la valeur sélective de son porteur ou si cette mutation est neutre sélectivement, elle sera éliminée à plus ou moins longue échéance.Ces considérations suggèrent que la phase la plus récente d’un polymorphisme doit être favorable à l’organisme pour pouvoir atteindre une fréquence supérieure à celle qui est produite par la seule mutation récurrente (Ford, 1965). Il s’ensuit que l’étude des polymorphismes est intimement liée aux facteurs de sélection naturelle opérant dans les populations. Comme la sélection naturelle est avant tout un phénomène écologique dû à la compétition ou à la concurrence entre organismes (C. Darwin, 1859), le champ d’activités de ceux que préoccupent les problèmes de polymorphismes permet d’effectuer une jonction entre la génétique traditionnelle des populations et l’écologie. L’importance de l’étude du polymorphisme est soulignée par J. L. Hubby et R. C. Lewontin (1966), lorsqu’ils disent qu’une «description de la variation génétique existant dans une population constitue la donnée fondamentale des travaux sur l’évolution» [cf. GÉNÉTIQUE]. Dans cet article, cette description sera abordée sous l’angle quantitatif au niveau allélique avant d’être mise en corrélation avec des facteurs écologiques au niveau de la population ou de l’espèce.L’aspect quantitatif du polymorphismeLa structure d’une population peut être définie en termes de fréquences géniques (cf. génétique des POPULATIONS). Les gènes étant présents sous la forme d’un couple d’allèles dans chaque individu diploïde, les individus d’une population sont soit homozygotes (les deux allèles étant semblables), soit hétérozygotes (les deux allèles étant différents) à chaque locus considéré. Il n’y a polymorphisme que si une population donnée contient au moins un individu hétérozygote à un locus donné. Pour quantifier le polymorphisme, il est nécessaire alors de déterminer la proportion de loci hétérozygotes dans le génome d’un individu ou encore la proportion de loci hétérozygotes dans le pool de gènes d’une population.Le problème ainsi posé paraît simple. Pour y répondre ne suffirait-il pas, en effet, d’établir la liste des loci connus pour telle ou telle population et de déterminer ensuite ceux qui possèdent des allèles distincts? Ces informations permettraient de calculer la proportion de loci hétérozygotes et, partant, d’estimer le pourcentage de ces derniers dans l’ensemble du génome ou du pool de gènes. En fait, comme l’a souligné Lewontin (1967), il faut des critères solides pour pouvoir discriminer les allèles et disposer donc d’une méthodologie suffisante. Or les méthodes classiques de la génétique des populations ne sont pas adéquates à l’obtention de données précises au niveau allélique. Ces méthodes portent soit sur l’analyse de polymorphismes décelables par examen visuel de phénotypes morphologiques (par exemple le polymorphisme de l’escargot Cepaea ), soit sur l’étude de polymorphismes «cachés», représentés par des mutants, dont les manifestations phénotypiques se traduisent par des effets délétères, allant jusqu’à la létalité. Dans le premier cas, il est rare que l’on connaisse à fond le mode héréditaire de ces polymorphismes. Lorsque cette connaissance existe, comme pour Cepaea , on s’aperçoit que l’on n’a pas affaire à des systèmes alléliques simples, mais à des séries de loci, chacun ayant des allèles multiples, l’ensemble étant rendu plus compliqué encore par des phénomènes de liaison génétique. Dans le second cas, celui des polymorphismes cachés, il est possible de mettre en évidence quantitativement l’effet collectif de ces variations par des techniques complexes qui permettent l’accumulation de mutations sur certains chromosomes. Mais, ici, cet effet collectif se traduit par une dépression de la viabilité des souches expérimentales au niveau de la population tout entière et non au niveau allélique dans chaque individu, et l’analyse détaillée suggère que plusieurs centaines de loci y participent.Hubby et Lewontin (1966) ont fort bien exprimé les raisons de l’échec des méthodes conventionnelles. Pour eux, les quatre conditions suivantes devraient être remplies:– Des différences phénotypiques dues à des allèles distincts doivent être observées à chaque locus et pour chaque individu.– Les allèles d’un locus donné doivent être distingués des allèles des autres loci.– Les allèles d’un locus donné doivent être distingués les uns des autres.– Les loci qui sont recensés doivent constituer un échantillonnage représentatif du génome quant à leurs effets phénotypiques.L’application de l’électrophorèse à la génétique des populations a permis d’obtenir des résultats suffisamment précis pour pouvoir répondre à la question posée plus haut, car les quatre critères ci-dessus sont bien davantage respectés par cette technique que par les méthodes classiques.Polymorphisme biochimique et hétérozygotie géniqueDepuis 1965 environ, l’électrophorèse [cf. ÉLECTROPHORÈSE] sur gels d’amidon ou de polyacrylamide a été utilisée pour mesurer l’hétérozygotie génique d’individus et de populations de nombreux organismes, allant des protozoaires aux vertébrés, en passant par les plantes vasculaires et les invertébrés.Les protéines d’un organisme constituent un excellent objet pour analyser les polymorphismes au niveau génique [cf. PROTÉINES]. Chaque protéine est composée d’un arrangement linéaire d’acides aminés formant des séquences déterminées directement par les gènes (fraction d’ADN). Beaucoup d’acides aminés ont des chaînes latérales, chargées électriquement, qui s’écartent de l’axe de la protéine et lui confèrent sa charge totale nette. On peut séparer des protéines chargées différemment en les faisant migrer dans un champ électrique. Une mutation, en particulier la substitution d’un acide aminé par un autre dans la molécule d’ADN, qui aboutit à un changement de charge, peut être décélée par une modification de la mobilité électrophorétique lorsque l’on compare la migration d’une protéine mutée avec celle d’une non mutée. Ainsi la variabilité génétique d’une population peut être mesurée en termes de mobilités électrophorétiques d’une large gamme de protéines extraites d’un échantillon de cette population.Le milieu dans lequel les protéines migrent et où l’on introduit le champ électrique est le plus souvent un gel à base d’amidon ou de polyacrylamide. On place un échantillon de protéine, provenant d’un tissu donné (sang, foie, etc.) préalablement traité, sur le milieu de soutien (gel), saturé de solution tampon et qui est en contact avec des électrodes à chaque extrémité. Lorsque l’on introduit une différence de voltage, le courant passe à travers le gel et les protéines sont entraînées dans un mouvement de migration. Après ce déplacement, les différentes fractions protéiniques sont mises en évidence par des colorants histochimiques appropriés à des protéines spécifiques (enzymes) ou à des groupes de protéines. Dans de nombreux cas, plusieurs bandes apparaissent alors sur le gel. Ces bandes indiquent la présence de plusieurs protéines ayant chacune une mobilité distincte et pouvant catalyser une réaction biochimique particulière. Ces variantes d’une enzyme sont appelées isozymes. En principe, les isozymes peuvent avoir plusieurs origines, et notamment, ce qui nous intéresse ici, des variantes produites par les allèles d’un seul locus génétique qu’on appelle alors allozymes. L’analyse génétique du mode d’hérédité d’un assez grand nombre d’isozymes, chez des mouches, des moustiques, des souris, des tourterelles et chez l’homme entre autres, a permis de démontrer sans équivoque possible que ces variantes sont bel et bien des allozymes et se comportent suivant les lois de ségrégation mendéliennes.La lecture de tels gels traités par électrophorèse et coloration histochimique permet à l’investigateur de dire combien de protéines possèdent des variantes et combien n’en possèdent pas, combien d’individus sont homozygotes ou hétérozygotes, et de dénombrer les loci possédant des allèles distincts. De cette manière, il est possible d’estimer la proportion de loci hétérozygotes par individu ou par population.Combien de loci sont-ils polymorphiques?Les premières réponses à cette question ont été apportées par des travaux sur des organismes aussi divers que des invertébrés marins, des grenouilles, des souris, des drosophiles et l’homme. On a ainsi trouvé que 6 à 12 p. 100 des loci sont hétérozygotes par individu et que 14 à 42 p. 100 sont hétérozygotes par population (Selander, Yang, Lewontin et Johnson, 1970). Il y a donc une proportion très importante de loci polymorphiques, que ce soit au niveau de l’individu ou au niveau de la population. Comme ordre de grandeur moyen, on peut dire que chaque individu a des allèles différents à un locus sur dix et que chaque population est polymorphe à un locus sur trois dans le pool de gènes. Si l’on admet que l’homme possède environ 100 000 loci dans son génome, 10 000 de ces loci sont polymorphes. Il y a donc énormément d’hétérozygotie dans les populations: c’est là le fait majeur apporté par les recherches récentes. Mais les choses ne sont bien entendu pas si simples que cela. De nombreuses études effectuées depuis lors ont démontré que la variabilité génétique, révélée par les analyses électrophorétiques, n’est pas présente à des proportions plus ou moins constantes dans chaque individu, dans chaque population ou dans chaque espèce, mais que ces proportions varient beaucoup, souvent en parallèle avec certains facteurs écologiques ou géographiques. On peut envisager l’étude de cette diversité d’abord au niveau interspécifique, pour tenter d’établir des rapports avec les processus évolutifs de spéciation (formation de nouvelles espèces), et ensuite au niveau intraspécifique, pour tenter de discerner les modalités des polymorphismes en termes de sélection naturelle.Variation interspécifiqueLa spéciation dépend de la présence, dans des isolats géographiques, d’une variabilité génétique suffisante pour que la sélection puisse agir. Une fois le processus de formation d’espèces nouvelles terminé, et si les deux espèces récemment formées ont des aires géographiques chevauchantes, elles présentent presque toujours des différences morphologiques qui sous-entendent des polymorphismes étendus et différents dans chaque espèce. Ces différences sont en général aisément démontrées qualitativement, mais il est très difficile de les quantifier. En 1967, ce problème était pour Lewontin «le plus important problème sans solution en génétique des populations».Dès 1984, trois groupes d’organismes ont été particulièrement bien étudiés: les drosophiles, certains lézards et plusieurs groupes de rongeurs, pour lesquels on possède des informations sur les ressemblances ou les différences entre espèces au niveau des polymorphismes géniques. Ces données ont en général été analysées par des coefficients de similarité génétique, par exemple:
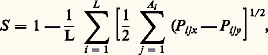 formule où la similarité génétique S est donnée par L (le nombre de loci recensés), par Ai (le nombre d’allèles au locus i ) et par Pijx et Pijy (les fréquences de l’allèle j au locus i dans les populations x et y ). Utilisant S pour comparer des espèces deux à deux pour L loci et A allèles à n loci, on peut alors obtenir des estimations sur la manière dont ces espèces diffèrent les unes des autres. En d’autres termes, au lieu de dire: l’espèce A ressemble davantage à B qu’à C parce qu’elle est plus ou moins grande, plus ou moins pigmentée, etc., on peut dire : A ressemble davantage à B qu’à C parce que le coefficient S indique x pour cent de similarité génétique, etc. Le tableau 1 résume certaines données sur les valeurs de S chez des drosophiles, des lézards et des rongeurs. Que suggèrent ces données?En tout premier lieu, elles suggèrent qu’il y a une marge de variation considérable dans le degré de similarité génétique entre espèces. Ainsi les quatre espèces du genre Anolis sont très peu semblables les unes aux autres (S est petit), tandis que les espèces du genre Drosophila ainsi que les espèces de rongeurs des genres Dipodomys et Sigmodon sont très semblables (S est grand). Mais, parmi les rongeurs, il existe aussi de grandes différences dans les valeurs de S d’un genre à l’autre.Si l’on examine maintenant le problème sous l’angle du nombre de protéines que possèdent en commun les espèces congénériques de ces divers groupes, on obtient une autre série de résultats, en particulier en étudiant les espèces jumelles, c’est-à-dire celles qui sont très proches parentes évolutivement et qui sont extrêmement semblables par leur morphologie. Chez Drosophila , les espèces jumelles possèdent en commun entre 13 et 86 p. 100 de leurs protéines ayant une même mobilité électrophorétique. Il y a donc une très grande variabilité lorsque l’on considère seulement les extrêmes, bien que la moyenne semble osciller autour de 50 p. 100, ce qui suggère la conclusion qualitative que les espèces jumelles de drosophiles sont bien différenciées génétiquement, car elles ne se partagent que la moitié de leurs protéines. Chez Anolis , les données sur les espèces non jumelles sont les suivantes: en moyenne, elles ont en commun 20 p. 100 des protéines, alors que des sous-espèces en ont environ 56 p. 100 en commun. Chez les Dipodomys , dont les espèces ne sont pas jumelles, les valeurs de S sont proches de celles de Drosophila (tabl. 1).À première vue, ces résultats sont décevants. Malgré une apparence de quantification, il ne semble pas possible de généraliser et l’on est ramené à des estimations qualitatives, différentes pour chaque groupe; il serait donc nécessaire d’expliquer chaque cas par une hypothèse ad hoc . Cependant, les résultats résumés plus haut montrent clairement que les espèces, qu’elles soient jumelles ou non, diffèrent entre elles par une fraction importante de leurs loci, si l’on en juge par l’échantillonnage de ces derniers. Une différence de l’ordre de 10 p. 100 seulement représente des différences au niveau génique se chiffrant par centaines ou par milliers de loci. Bien qu’il ne soit pas possible de déterminer exactement le nombre de loci différents, il est clair que le phénomène de spéciation s’accompagne d’une réorganisation des pools de gènes des espèces naissantes (F. J. Ayala, 1975).Variation intraspécifiqueEn plus des trois groupes – mouches, lézards et rongeurs – cités plus haut, beaucoup d’autres organismes ont été étudiés sous l’angle de la variation intraspécifique des niveaux de polymorphisme. On dispose par conséquent d’une documentation importante. Un premier essai de synthèse a été publié par C. Manwell et C. M. A. Baker (1970), et d’autres ont suivi depuis lors (G. Pasteur, 1974, J. R. Powell, 1975; J. C. Avise et C. F. Aquadro, 1982). Les réflexions qui suivent représentent un échantillonnage d’une littérature abondante sur des groupes allant des vertébrés aux invertébrés et des plantes aux animaux; ces commentaires sont rédigés sous la forme de généralisations, qui toutes souffrent d’exceptions mais qui peuvent servir d’hypothèses.1. Les populations occupant la région centrale de l’aire géographique de l’espèce dont elles font partie sont plus variables que les populations marginales. En d’autres termes, elles ont un degré plus élevé d’hétérozygotie, ce qui se traduit par un plus fort pourcentage de leurs loci hétérozygotes par individu ou par population.2. Les populations qui sont géographiquement isolées par une barrière écogéographique quelconque, par exemple les populations insulaires d’une espèce continentale, ont moins d’hétérozygotie que les populations continentales. Les propositions 1 et 2 sont illustrées clairement par les données du tableau 2 sur Drosophila pseudoobscura .3. La variation géographique de fréquences d’allèles à de nombreux loci biochimiques est en corrélation avec des variations graduelles de certains paramètres du milieu, en particulier la température de l’eau chez plusieurs organismes aquatiques, marins ou d’eau douce, comme les mollusques ou les poissons.4. On retrouve souvent les mêmes allèles, et ce qui est plus intéressant à des fréquences très semblables, dans des populations très éloignées géographiquement les unes des autres ou à des périodes de l’année différentes, donc pendant des phases distinctes du cycle annuel. Il y a par conséquent une stabilité apparente dans la distribution de certains polymorphismes protéiniques dans les deux dimensions écologiques du temps et de l’espace. Bien entendu, le contraire est vrai aussi, mais pour d’autres loci: il y a une variation parallèle à des facteurs écogéographiques (cf. supra ).5. Des milieux, hétérogènes dans le temps ou l’espace, abritent en général des populations dont le degré d’hétérozygotie est plus élevé que celui des populations habitant des milieux plus homogènes. Cela est vrai non seulement dans la nature, lorsque l’on compare des populations d’une même espèce occupant des milieux relativement stables (grottes) avec des populations vivant en milieu plus variable (surface), mais aussi en laboratoire où l’on peut reproduire de manière simplifiée certains types de variabilité des milieux. Le tableau 3 illustre une telle situation étudiée chez Drosophila willistoni . Un autre moyen de confirmer cette observation est de comparer plusieurs espèces d’un même groupe taxonomique qui vivent le long d’un gradient écologique représentant la stabilité relative du milieu. On note alors que les espèces vivant à l’extrémité plus stable du gradient ont plus d’allèles par locus que celles qui vivent à l’autre extrémité, moins stable. Par stabilité il faut entendre ici la régularité relative des fluctuations physico-chimiques et biotiques du milieu. Un exemple étudié chez des bivalves est la répartition des espèces en fonction de la profondeur du substrat (boues meubles) dans la zone tidale et subtidale. Plus l’espèce s’enfouit dans le substrat, plus les loci sont polymorphes, et moins grandes sont les variations du milieu. Il y a toutefois une exception notable: les espèces d’invertébrés marins des grands fonds (plus de 1 000 mètres) analysées à ce jour, et qui vivent par conséquent dans un milieu très stable, présentent tout autant de polymorphismes que d’autres espèces vivant dans des milieux bien moins stables, dans la zone tidale.6. Finalement, il semble que des espèces appartenant à des lignées phylogénétiquement très anciennes (comme Limulus ) possèdent tout autant d’hétérozygotie que des espèces appartenant à des lignées beaucoup plus récentes et en pleine évolution (comme Mus ).Les données citées plus haut au sujet du polymorphisme au sein de nombreuses populations d’espèces fort diverses suggèrent très clairement qu’il y a une série de corrélations complexes entre certains facteurs écologiques du milieu ambiant et le genre de variabilité des niveaux d’hétérozygotie. On pourrait donc conclure que ces corrélations traduisent l’influence de la sélection naturelle, dont les modalités d’action dépendent précisément du type de milieu. Cette conclusion est celle à laquelle ont abouti de nombreux auteurs, mais il n’en reste pas moins que l’étude exacte de ces modalités sélectives constitue un champ de recherche encore relativement peu exploré. Soit sur le terrain, soit en laboratoire, un certain nombre de travaux ont déjà été effectués sur des organismes variés, allant des plantes vasculaires aux vertébrés, et ont déjà démontré l’extrême complication de l’action sélective. En dépit de ces démonstrations, plusieurs généticiens des populations ont avancé des arguments en faveur de l’hypothèse que les très nombreux polymorphismes biochimiques ne sont pas maintenus par la sélection naturelle.Le maintien du polymorphismeLewontin et Hubby (1966) ont indiqué comment l’on peut envisager le problème du maintien du polymorphisme. La variation génétique est érodée par les phénomènes de dérive génétique dans des populations dont l’effectif est périodiquement diminué, ainsi que par la sélection contre les gènes délétères récessifs ou partiellement dominants. Par contre, la variation génétique est maintenue par la mutation, la migration génique et la sélection balancée ou diversifiante qui favorise les hétérozygotes. La variation décelée par électrophorèse pourrait alors être due à l’une des trois raisons suivantes:– les allèles sont neutres du point de vue sélectif;– la sélection élimine les allèles, mais la mutation les remplace;– la sélection favorise les hétérozygotes (hétérosis).Comme la deuxième raison sous-entend une pression mutationnelle bien au-delà de ce que l’on connaît actuellement, on peut l’éliminer. Les études faites à ce jour indiquent qu’il y a hétérosis mais pas neutralité sélective. Cependant, une hétérosis portant sur des milliers de loci aurait comme résultat un fardeau génétique tel qu’il semble douteux que la population puisse survivre. Il y a donc un dilemme, et c’est précisément pour cela que certains, comme M. Kimura et T. Ohta (1971), soutiennent l’hypothèse de neutralité sélective. Si un allèle mutant est neutre sélectivement, son élimination fortuite de la population aura une probabilité de 1 – 1/2 N . Dans ce cas, le temps nécessaire à l’élimination sera de:
formule où la similarité génétique S est donnée par L (le nombre de loci recensés), par Ai (le nombre d’allèles au locus i ) et par Pijx et Pijy (les fréquences de l’allèle j au locus i dans les populations x et y ). Utilisant S pour comparer des espèces deux à deux pour L loci et A allèles à n loci, on peut alors obtenir des estimations sur la manière dont ces espèces diffèrent les unes des autres. En d’autres termes, au lieu de dire: l’espèce A ressemble davantage à B qu’à C parce qu’elle est plus ou moins grande, plus ou moins pigmentée, etc., on peut dire : A ressemble davantage à B qu’à C parce que le coefficient S indique x pour cent de similarité génétique, etc. Le tableau 1 résume certaines données sur les valeurs de S chez des drosophiles, des lézards et des rongeurs. Que suggèrent ces données?En tout premier lieu, elles suggèrent qu’il y a une marge de variation considérable dans le degré de similarité génétique entre espèces. Ainsi les quatre espèces du genre Anolis sont très peu semblables les unes aux autres (S est petit), tandis que les espèces du genre Drosophila ainsi que les espèces de rongeurs des genres Dipodomys et Sigmodon sont très semblables (S est grand). Mais, parmi les rongeurs, il existe aussi de grandes différences dans les valeurs de S d’un genre à l’autre.Si l’on examine maintenant le problème sous l’angle du nombre de protéines que possèdent en commun les espèces congénériques de ces divers groupes, on obtient une autre série de résultats, en particulier en étudiant les espèces jumelles, c’est-à-dire celles qui sont très proches parentes évolutivement et qui sont extrêmement semblables par leur morphologie. Chez Drosophila , les espèces jumelles possèdent en commun entre 13 et 86 p. 100 de leurs protéines ayant une même mobilité électrophorétique. Il y a donc une très grande variabilité lorsque l’on considère seulement les extrêmes, bien que la moyenne semble osciller autour de 50 p. 100, ce qui suggère la conclusion qualitative que les espèces jumelles de drosophiles sont bien différenciées génétiquement, car elles ne se partagent que la moitié de leurs protéines. Chez Anolis , les données sur les espèces non jumelles sont les suivantes: en moyenne, elles ont en commun 20 p. 100 des protéines, alors que des sous-espèces en ont environ 56 p. 100 en commun. Chez les Dipodomys , dont les espèces ne sont pas jumelles, les valeurs de S sont proches de celles de Drosophila (tabl. 1).À première vue, ces résultats sont décevants. Malgré une apparence de quantification, il ne semble pas possible de généraliser et l’on est ramené à des estimations qualitatives, différentes pour chaque groupe; il serait donc nécessaire d’expliquer chaque cas par une hypothèse ad hoc . Cependant, les résultats résumés plus haut montrent clairement que les espèces, qu’elles soient jumelles ou non, diffèrent entre elles par une fraction importante de leurs loci, si l’on en juge par l’échantillonnage de ces derniers. Une différence de l’ordre de 10 p. 100 seulement représente des différences au niveau génique se chiffrant par centaines ou par milliers de loci. Bien qu’il ne soit pas possible de déterminer exactement le nombre de loci différents, il est clair que le phénomène de spéciation s’accompagne d’une réorganisation des pools de gènes des espèces naissantes (F. J. Ayala, 1975).Variation intraspécifiqueEn plus des trois groupes – mouches, lézards et rongeurs – cités plus haut, beaucoup d’autres organismes ont été étudiés sous l’angle de la variation intraspécifique des niveaux de polymorphisme. On dispose par conséquent d’une documentation importante. Un premier essai de synthèse a été publié par C. Manwell et C. M. A. Baker (1970), et d’autres ont suivi depuis lors (G. Pasteur, 1974, J. R. Powell, 1975; J. C. Avise et C. F. Aquadro, 1982). Les réflexions qui suivent représentent un échantillonnage d’une littérature abondante sur des groupes allant des vertébrés aux invertébrés et des plantes aux animaux; ces commentaires sont rédigés sous la forme de généralisations, qui toutes souffrent d’exceptions mais qui peuvent servir d’hypothèses.1. Les populations occupant la région centrale de l’aire géographique de l’espèce dont elles font partie sont plus variables que les populations marginales. En d’autres termes, elles ont un degré plus élevé d’hétérozygotie, ce qui se traduit par un plus fort pourcentage de leurs loci hétérozygotes par individu ou par population.2. Les populations qui sont géographiquement isolées par une barrière écogéographique quelconque, par exemple les populations insulaires d’une espèce continentale, ont moins d’hétérozygotie que les populations continentales. Les propositions 1 et 2 sont illustrées clairement par les données du tableau 2 sur Drosophila pseudoobscura .3. La variation géographique de fréquences d’allèles à de nombreux loci biochimiques est en corrélation avec des variations graduelles de certains paramètres du milieu, en particulier la température de l’eau chez plusieurs organismes aquatiques, marins ou d’eau douce, comme les mollusques ou les poissons.4. On retrouve souvent les mêmes allèles, et ce qui est plus intéressant à des fréquences très semblables, dans des populations très éloignées géographiquement les unes des autres ou à des périodes de l’année différentes, donc pendant des phases distinctes du cycle annuel. Il y a par conséquent une stabilité apparente dans la distribution de certains polymorphismes protéiniques dans les deux dimensions écologiques du temps et de l’espace. Bien entendu, le contraire est vrai aussi, mais pour d’autres loci: il y a une variation parallèle à des facteurs écogéographiques (cf. supra ).5. Des milieux, hétérogènes dans le temps ou l’espace, abritent en général des populations dont le degré d’hétérozygotie est plus élevé que celui des populations habitant des milieux plus homogènes. Cela est vrai non seulement dans la nature, lorsque l’on compare des populations d’une même espèce occupant des milieux relativement stables (grottes) avec des populations vivant en milieu plus variable (surface), mais aussi en laboratoire où l’on peut reproduire de manière simplifiée certains types de variabilité des milieux. Le tableau 3 illustre une telle situation étudiée chez Drosophila willistoni . Un autre moyen de confirmer cette observation est de comparer plusieurs espèces d’un même groupe taxonomique qui vivent le long d’un gradient écologique représentant la stabilité relative du milieu. On note alors que les espèces vivant à l’extrémité plus stable du gradient ont plus d’allèles par locus que celles qui vivent à l’autre extrémité, moins stable. Par stabilité il faut entendre ici la régularité relative des fluctuations physico-chimiques et biotiques du milieu. Un exemple étudié chez des bivalves est la répartition des espèces en fonction de la profondeur du substrat (boues meubles) dans la zone tidale et subtidale. Plus l’espèce s’enfouit dans le substrat, plus les loci sont polymorphes, et moins grandes sont les variations du milieu. Il y a toutefois une exception notable: les espèces d’invertébrés marins des grands fonds (plus de 1 000 mètres) analysées à ce jour, et qui vivent par conséquent dans un milieu très stable, présentent tout autant de polymorphismes que d’autres espèces vivant dans des milieux bien moins stables, dans la zone tidale.6. Finalement, il semble que des espèces appartenant à des lignées phylogénétiquement très anciennes (comme Limulus ) possèdent tout autant d’hétérozygotie que des espèces appartenant à des lignées beaucoup plus récentes et en pleine évolution (comme Mus ).Les données citées plus haut au sujet du polymorphisme au sein de nombreuses populations d’espèces fort diverses suggèrent très clairement qu’il y a une série de corrélations complexes entre certains facteurs écologiques du milieu ambiant et le genre de variabilité des niveaux d’hétérozygotie. On pourrait donc conclure que ces corrélations traduisent l’influence de la sélection naturelle, dont les modalités d’action dépendent précisément du type de milieu. Cette conclusion est celle à laquelle ont abouti de nombreux auteurs, mais il n’en reste pas moins que l’étude exacte de ces modalités sélectives constitue un champ de recherche encore relativement peu exploré. Soit sur le terrain, soit en laboratoire, un certain nombre de travaux ont déjà été effectués sur des organismes variés, allant des plantes vasculaires aux vertébrés, et ont déjà démontré l’extrême complication de l’action sélective. En dépit de ces démonstrations, plusieurs généticiens des populations ont avancé des arguments en faveur de l’hypothèse que les très nombreux polymorphismes biochimiques ne sont pas maintenus par la sélection naturelle.Le maintien du polymorphismeLewontin et Hubby (1966) ont indiqué comment l’on peut envisager le problème du maintien du polymorphisme. La variation génétique est érodée par les phénomènes de dérive génétique dans des populations dont l’effectif est périodiquement diminué, ainsi que par la sélection contre les gènes délétères récessifs ou partiellement dominants. Par contre, la variation génétique est maintenue par la mutation, la migration génique et la sélection balancée ou diversifiante qui favorise les hétérozygotes. La variation décelée par électrophorèse pourrait alors être due à l’une des trois raisons suivantes:– les allèles sont neutres du point de vue sélectif;– la sélection élimine les allèles, mais la mutation les remplace;– la sélection favorise les hétérozygotes (hétérosis).Comme la deuxième raison sous-entend une pression mutationnelle bien au-delà de ce que l’on connaît actuellement, on peut l’éliminer. Les études faites à ce jour indiquent qu’il y a hétérosis mais pas neutralité sélective. Cependant, une hétérosis portant sur des milliers de loci aurait comme résultat un fardeau génétique tel qu’il semble douteux que la population puisse survivre. Il y a donc un dilemme, et c’est précisément pour cela que certains, comme M. Kimura et T. Ohta (1971), soutiennent l’hypothèse de neutralité sélective. Si un allèle mutant est neutre sélectivement, son élimination fortuite de la population aura une probabilité de 1 – 1/2 N . Dans ce cas, le temps nécessaire à l’élimination sera de: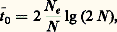 e étant la population mendélienne effective. Le temps jusqu’à fixation sera de:
e étant la population mendélienne effective. Le temps jusqu’à fixation sera de: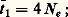 il y aura alors de nombreux polymorphismes à un grand nombre de loci et les allèles seront maintenus entre élimination et fixation (dues toutes deux à des facteurs purement aléatoires) avant d’être périodiquement «remis» dans la population par mutation. Théoriquement, ce point de vue paraît raisonnable, mais pratiquement il ignore aveuglément la masse de données suggérant que les polymorphismes biochimiques ne sont pas neutres. Comme H. Harris (1971) l’a noté, l’hypothèse de neutralité a le mérite de remettre en question la solution d’un problème d’une extrême importance, le polymorphisme étant, de l’avis général, la base même de l’évolution.B. Clarke (1973) a clairement montré la direction à suivre à l’avenir, en disant qu’il fallait maintenant s’attaquer au problème des effets biochimiques des variantes enzymatiques pour découvrir si les différences décelées dans des cas précis sont ou non soumises à l’action de processus de sélection.
il y aura alors de nombreux polymorphismes à un grand nombre de loci et les allèles seront maintenus entre élimination et fixation (dues toutes deux à des facteurs purement aléatoires) avant d’être périodiquement «remis» dans la population par mutation. Théoriquement, ce point de vue paraît raisonnable, mais pratiquement il ignore aveuglément la masse de données suggérant que les polymorphismes biochimiques ne sont pas neutres. Comme H. Harris (1971) l’a noté, l’hypothèse de neutralité a le mérite de remettre en question la solution d’un problème d’une extrême importance, le polymorphisme étant, de l’avis général, la base même de l’évolution.B. Clarke (1973) a clairement montré la direction à suivre à l’avenir, en disant qu’il fallait maintenant s’attaquer au problème des effets biochimiques des variantes enzymatiques pour découvrir si les différences décelées dans des cas précis sont ou non soumises à l’action de processus de sélection.
Encyclopédie Universelle. 2012.